抗体是一种免疫球蛋白,由B淋巴细胞产生。所有的抗体分子都有相似的结构,都是由两条相同的重链(heavy chain,H链)和两条相同的轻链(light chain,L链)组成的4条肽链对称结构。轻、重链的链内和链间分别借助二硫键相连。如下为抗体的分子结构图:
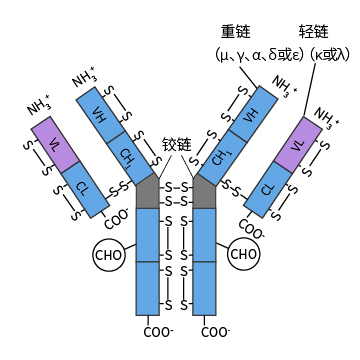
轻链
大约由214个氨基酸残基组成,通常不含碳水化合物,分子量约为24kDa。每条轻链含有两个由链内二硫键所组成的环肽。L链可分为κ与λ型,一个抗体分子中只能具有两型轻链的一种,在不同的物种中κ/λ比例不同。如小鼠抗体的κ:λ为20:1,而正常人抗体中κ:λ约为2:1。
重链
重链大小约为轻链的2倍,含450~550个氨基酸残基,分子量约为55或75kDa。根据H链抗原性的差异可将其分为5类:μ链、γ链、α链、δ链和ε链。不同H链与L 链(κ或λ链)组成完整Ig的分子分别称之为IgM、IgG、IgA、IgD和IgE。(抗体分类和亚型)
可变区(V区)
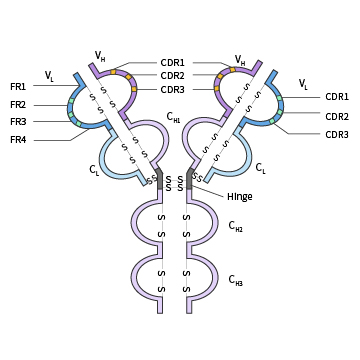
是抗体分子N端存在一段氨基酸序列变化较大的区域,由N端重链1/4和轻链1/2组成。可变区存在可以与抗原特异性结合的部位,即抗原结合位点,用以决定抗体识别的特异性。一个抗体有两个抗原结合位点,可以同时结合两个抗原分子。
某些特定位置的氨基酸残基的组成和排列顺序高度可变,称为超变区(HVR)又称为抗原互补决定区(CDR),共同组成Ig的抗原识别部位。骨架区(FR)为于V区中除HVR以外的部位,此部分AA替换频率较低的部分,便于稳定HVR的结构。可变区中CDR和FR的组成方式为“FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4”。
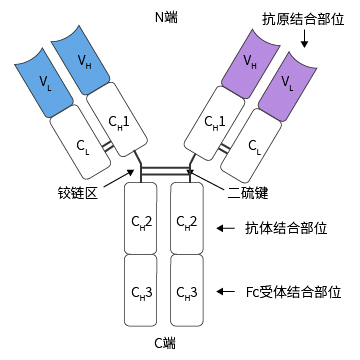
恒定区(C区)
抗体分子C端氨基酸相对稳定的区域,占轻链1/2和重链3/4或4/5。抗体轻链的恒定区由一个Ig结构域构成;重链的恒定区有3~4个串联的Ig结构域及一个用于增加灵活性的铰链区构成。主要作用于免疫应答,具有许多重要的生物学功能。
铰链区
在重链CH1尾部和CH2头部,有一个约30个氨基酸残基组成的柔性的铰链区(hinge region),其中脯氨酸含量极高,不形成α螺旋,易伸展弯曲,存在2~5个链间二硫键。
铰链区的主要功能是调节抗体可变区上抗原结合部位与相应的抗原表位匹配,促进抗原抗体结合,此外也利于Ig分子构象变化,暴露补体结合位点。对木瓜蛋白酶、胃蛋白酶敏感,当用这些蛋白酶水解免疫球蛋白分子时常此区发生裂解。IgM和IgE缺乏铰链区。
功能区
Ig分子的H链与L链可通过链内二硫键折叠成若干球形功能区,每一功能区(domain)约由110个氨基酸组成。在功能区中氨基酸序列有高度同源性。
- L链功能区分为L链可变区(VL)和L链恒定区(CL)两功能区。
- H链功能区IgG、IgA和IgD的H链各有一个可变区(VH)和三个恒定区(CH1、CH2和CH3)共四个功能区;
IgM和IgE的H链各有一个可变区(VH)和四个恒定区(CH1、CH2、CH3和CH4)共五个功能区。
L链和H链中V区或C区每个功能区各形成一个免疫球蛋白折叠(immunoglobulin fold),每个Ig折叠含有两个大致平行、由二硫连接的β片层结构(betapleated sheets),每个β片层结构由3至5股反平行的多肽链组成。可变区中的高变区在Ig折叠的一侧形成高变区环(hypervariable loops),是与抗原结合的位置。
作用
- VL和VH是与抗原结合的部位,其中HVR(CDR)是V区中与抗原决定簇(或表位)互补结合的部位。VH和VL通过非共价相互作用,组成一个FV区。单位Ig分子具有2个抗原结合位点(antigen-binding site),二聚体分泌型IgA具有4个抗原结合位点,五聚体IgM可有10个抗原结合位点。
- CL和CH上具有部分同种异型的遗传标记。
- IgG 中CH2具有补体Clq结合点,能活化补体的经典活化途径。母体IgG借助CH2部分可通过胎盘主动传递到胎体内。
- IgG中 CH3具有结合单核细胞、巨噬细胞、粒细胞、B细胞和NK细胞Fc段受体的功能。
Fab片段
IgG分子在木瓜蛋白酶的作用下可以被降解为两个Fab段和一个Fc段。Fab段有抗体轻链的可变区、轻链恒定区、重链的可变区及恒定区构成。可变区是与抗原结合的部位,因此Fab段又称为抗原结合段。
Fc段
Fc段包含了所有抗体分子共有的蛋白质序列以及各个类别独有的决定簇。Fc段有多种生物学活性,具有结合补体、结合Fc受体、通过胎盘等作用。
推荐阅读: 抗体亲和力成熟测定